Decision Making
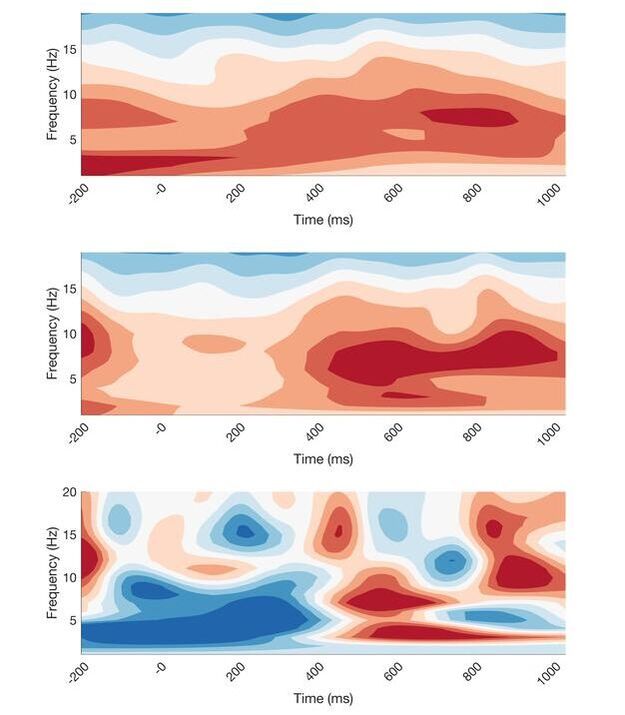
These are some preliminary results from my thesis study: "Portable EEG Measurement of Clinical Decision Making". In this experiment, participants were learning to diagnose rare and typical cases of liver decise. The idea here is to encourage analytical, System 2 thinking to solve rare cases, and measure this with frontal Theta EEG activity. This figure shows time-freqency wavelets for typical and rare cases, and the difference between the two. In the third wavelet, there seems to be a visual theta burst at around 500ms after stiumulus, indicating the engagement of System 2. These results are preliminary, and descriptive statistics have not been calculated for this data set yet.
|
My first area of research in the TAN Lab involved neural decision making, specifically under umbrella of Dual Process theory. Dual Process theory indicates that human reasoning can either be fast and intuitive, called System 1 thinking, or slow and analytical, called System 2 thinking. Multiple theories have emerged to prove and counter this idea, including Single Process accounts where decision making occurs on a spectrum between intuitive and analytical. While many disciplines have tackled this idea, including cognitive psychology and neuroscience, there is no consensus on the existence of Dual Process thinking. A goal of our lab is to contribute to this debate. Specifically, my colleagues and I have attempted to apply electroencephalography (EEG) and functional near-infrared spectroscopy (fNIRS) to examine neural correlates of System 1 and 2 thinking. One promising method is examing frequency data from decision making experiments, where some have found that bursts of Theta (4-8Hz) and Beta (12.5-30Hz) correlate with System 2 processing. This finding has led to a surge of new applications, including clinical decision making.
Some recommended readings related to my work: Williams, Kappen, Hassall, Wright, and Krigolson (2019). Thinking theta and alpha: Mechanisms of intuitive and analytical reasoning. Pennycook, Fugelsang, and Koehler (2015). What makes us think? A three-stage dual-process model of analytic engagement. |
Reinforcement Learning
|
Another research focus of mine involves hierarchical reinforcement learning, specifically the medial-frontal learning system (Holroyd and Coles, 2002). We use event-related potentials (ERPs) obtained from EEG data to observe neural learning signals during learning tasks. These tasks often operate on tenets of reinforcement learning, where actions are modified based on the reward associated with outcomes. Personally, I have focused on what neural learning signals look like in large sample sizes in an effort examine behavioural components that may affect learning. For example, my 2020 presentation at the Cognitive Neuroscience Society annual meeting showed our findings for the effects of sleep on the reward positivity, an ERP correlate of the medial-frontal learning system. Future work will focus on relating thoeries of hiearchical reinforcement learning to cognitive control.
Some recommended readings related to my work: Krigolson, Pierce, Holroyd, and Tanaka (2008). Learning to Become an Expert: Reinforcement Learning and the Acquisition of Perceptual Expertise Botvinick, Niv, and Barto (2009)- Hierarchically organized behavior and its neural foundations: A reinforcement learning perspective. |
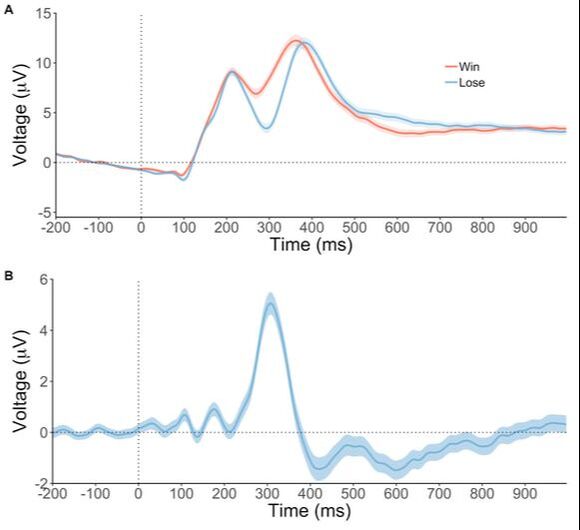
Here are ERP waveforms from 300 participants completing a two-armed bandit task. The task is designed to invoke the reward positivity, an ERP component that indexes feedback evaluation for the midbrain reinforcement learning system. The difference waveform presented here is a great example of a typical reward postitivity because of the large sample size. Shading represents 95% confidence intervals.
|
Cognitive Control
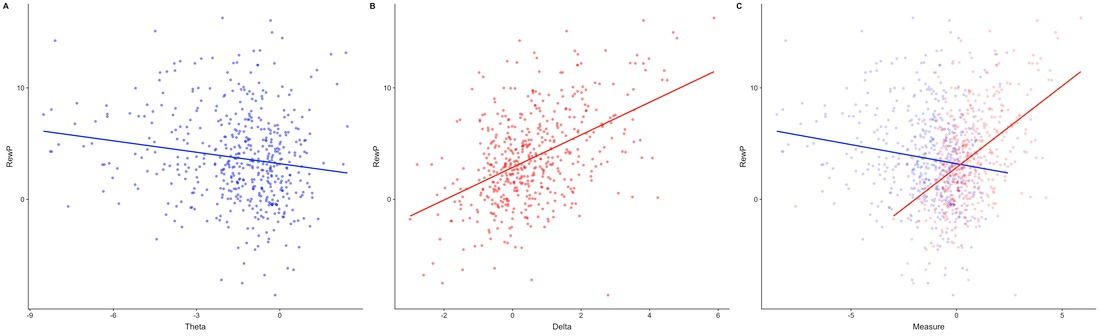
Here are some scatter plots showing reward positivity amplitude and frequency bands from the same 300 participant experiment I mentioned in the previous section. These results are interesting, especially the apparent negative correlation between the reward postivity amplitude and theta power. In future experiments, I plan to further explore where neural correlates of reinforcement learning and cognitive control work in various scenarios.
|
Cognitive control refers to the brain's ability to inhibit habitual mechanisms in favour of those that serve goal-directed action. For example, if I am making a difficult decision about where to eat dinner tonight, I may engage cognitive control to draw on long term memory of my favourite restaurants and decision-making systems to weigh the pros and cons of each restaurant, while shutting off systems unneccesary for the decision like auditory perception. Where I am beginning to direct focus is the intersection between cognitive control and hiearchical reinforcement learning, a field where many take a computational approach to where the two processes intersect for learning. Currently, I plan to conduct an experiment as part of my Master's thesis to explore the neural correlates of learning when working memory and reinforcement learning compete and cooperate throughout a decision.
Some recommended readings related to my work: Collins and Frank (2018). Within- and across-trial dynamics of human EEG reveal cooperative interplay between reinforcement learning and working memory. Colins and Frank (2016). Neural signature of hierarchically structured expectations predicts clustering and transfer of rule sets in reinforcement learning. |
Neural Basis of Teamwork
|
An area of neuroscience that has always held interest for me was the neural basis of teamwork between individuals. Many have found evidence for Interbrain Neural Synchrony (IBNS), the temporal and spatial coupling of neural oscillations between individuals cooperating with each other. IBNS research has lead to exploration on dynamics of large teams, emergence of leadership, and even the neural correlates of competition. Research in this area has huge implications for the workplace and education, where teamwork is being increasingly emphasized. I hope to start the process of this translation, as currently research is restricted to neuroimaging systems that are immobile and expensive. In the future, I will attempt to find a portable measure of IBNS by replicating established research tasks while neural data is recorded with various mobile EEG and fNIRS techniques. The aim of this research would be to create a system for actively measuring teamwork in applied settings that involve both cooperation and competition.
Some recommended readings related to my work: Dikker et al. (2017) Brain-to-Brain Synchrony Tracks Real-World Dynamic Group Interactions in the Classroom. Fishburn et al. (2018). Putting our heads together: interpersonal neural synchronization as a biological mechanism for shared intentionality. |
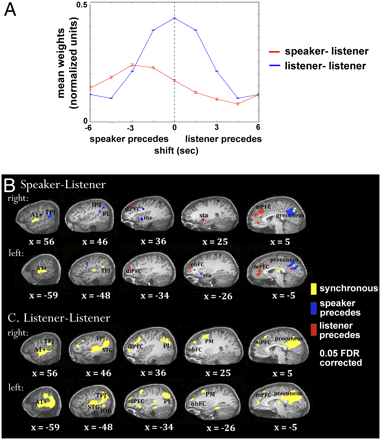
To highlight what IBNS looks like, here are results from Stephens et al. (2010)-"Speaker–listener neural coupling underlies successful communication". Their results were one of the early examples of IBNS that show the temporal and spatial dynamics of cooperation. As you can see, different brain areas for communication are activated earlier in the listener, earlier in the speaker, or at the same time for both. The key for my application is translating this baseline into more complex interactions involved with shared intention. This figure is particularly important to me because it was one of the first times I really dived into something out of my depth. I started reading about IBNS very soon after getting into the lab, and I knew absolutely nothing about fMRI. I became dedicated to learning fMRI so I could present my study ideas to my colleagues, which presented an oppurtunity to do self-driven learning about a completelty novel concept.
|
Want to read more about cognitive neuroscience topics?
I am currently curating a reading list for the TAN Lab that includes papers we all refer to often, or just have cool results. When I was starting my neuroscience career, I knew what I was interested in but had not idea where to start. My hope with this list is to help people in that exact situation. Check it out here!
